Morfologia
e Fisiologia
A palavra “morfologia” refere-se ao ramo da biologia
que lida com a forma e estrutura dos
organismos assim como à forma e estrutura de um organismo específico como um
todo.
A tabela MORFOLOGIA tem como objectivo preencher dois
objectivos que estão relacionados entre si:
i) Fornecer descrições concisas padronizadas e
portanto comparáveis de peixes incluídos nesta FishBase; e
As características
merísticas, morfométricas e descritivas identificam uma espécie
ii) Permitir uma identificação
rápida das espécies baseada nos caracteres usados em i).
Nos peixes, a maior parte das características usadas
na descrição e identificação são descritivas (quando se referem a caracteres
distintivos, ex. forma da barbatana caudal), morfométricos (quando se referem a
varáveis contínuas, ex. comprimento da cabeça como fracção do comprimento do
corpo) ou merísticas (quando se referem a variáveis descontínuas, ex. número de
raios ou espinhos na barbatana dorsal).
A tabela MORFOLOGIA incorpora características
descritivas em campos de escolha múltipla, e características morfométricas e
merísticas em campos numéricos. São principalmente as características
merísticas que são utilizadas para uma rápida identificação, seguida de
esquemas de identificação da base de dados de Froese e Papasissi (1990). A
estrutura da tabela MORFOLOGIA e dos seus campos de escolha é baseada em
estudos pormenorizados de grandes textos de ictiologia (ex. Lagler et al. 1977)
e de consultas a vários especialistas. Alguns termos aqui utilizados são muito
específicos; a sua definição pode ser encontrada no glossário da FishBase.
Campos A tabela MORFOLOGIA contém 67 campos de escolha, 79
campos numéricos e vários campos de notas.
Os campos de escolha dão ao utilizador opções
préprogramadas de descrições de partes de corpo ou características (ex. secção - circular, oval, comprimida,
achatada, angular, outra (ver Notas).
As escolhas incluídas foram reduzidas a um mínimo, e incluem apenas descrições
gerais e cobrem apenas as formas mais comuns. Na maior parte dos casos, a
escolha “outra” (ver Notas) é incluída para as espécies que possam ter formas
ou partes do corpo aberrantes. Quando “outra” é escolhida para um campo, uma
descrição detalhada da parte do corpo particular é incluída no campo Notas.
Por outro lado, os campos numéricos são utilizados
para as características morfométricas e merísticas. Na maior parte dos casos os
intervalos entram em campos separados de limites inferiores e superiores.
Quando um intervalo de vários valores é referido na literatura, mas o campo
permite apenas a entrada de um único valor (como nos campos proporção do
corpo), é a média dos valores disponíveis que entra.
O campo Notas integra características que, ou
não foram incluídas nos campos de escolha, ou precisam de descrições mais
detalhadas. Nesses campos, sublinha-se as características distintas e como
podem ser encontradas em espécies muito próximas. Notas sobre a variação da cor
(ontogénica, sexual ou geográfica) entram também, quando disponíveis, nestes
campos.
Devido
ao aumento do número de espécies na FishBase, achamos que seria um processo
moroso preencher os mais de 140 campos da tabela MORFOLOGIA para todas as
espécies. Foi decidido reduzir o número de campos activos aos normalmente
existentes em livros de taxonomia (diagnóstico e características merísticas
típicas) e preenchê-los regularmente. Esta tarefa foi completada entretanto
para todos os peixes ósseos do Japão e Colômbia Britânica e para todos os
peixes ósseos marinhos da Micronésia e África do Sul (Smith and Heemstra,
1986). Também se encontram completas todas as famílias abrangidas nos catálogos
FAO ou na série Indo-Pacific Fishes
de Randall. Tencionamos completar e verificar a tabela MORFOLOGIA por família
(Caixa 1, neste volume), utilizando referências faunísticas essenciais como Freshwater Fishes of Southern Africa
(Skelton, 1993).
Utilizações A utilização mais importante da informação contida na
tabela MORFOLOGIA, é uma rápida identificação do peixe (ver também a secção
IDENTIFICAÇÃO). O procedimento corrente préprogramado requer uma quantidade
mínima de informação como critério de procura:
·
Os dados da tabela MORFOLOGIA podem ser usados para
uma identificação rápida
Área da FAO na qual o peixe
foi recolhido
·
habitat (água doce,
salobra ou salgada)
·
Profundidade à qual o
peixe foi recolhido
·
Tamanho do especimen
·
nº de espinhos da
barbatana dorsal
·
nº de raios moles da
barbatana dorsal
·
nº de espinhos da
barbatana anal
·
nº de raios moles da
barbatana anal
·
A pesquisa, resulta geralmente em menos de 10
espécies possíveis
Ordem ou família taxonómica
(como opção)
Depois
procura através da base de dados e mostra a lista de espécies que coincidem com
os critérios fornecidos pelo utilizador. Normalmente, essa pesquisa resulta
numa lista de menos de 10 espécies da mesma família. O utilizador pode então
através das imagens e das descrições completas verificar a sua identificação.
Este
trabalho de procura também funciona com um ou mais campos de escolha deixados em
branco; nesses casos, a lista de espécies gerada torna-se mais longa. É de
notar no entanto que, até à data, a tabela MORFOLOGIA contém dados de apenas
5700 espécies. A informação disponível varia em grau de complexidade: muito
escassa, como no caso de Pellona
castelnaena; quase completa, como no caso de Lutjanus biguttatus. Saliente-se que os dados ainda não foram todos
verificados e por isso podem conter alguns erros.
Relatórios Procedimentos préprogramados para impressão de
Sinopses de Espécies e Sumários, utilizam informação da tabela MORFOLOGIA. O
procedimento para saída de um sumário de uma espécie, por exemplo, é extraído
do campo Características adicionais
(que dá descrições distintas), das contagens de elementos das barbatanas
dorsais e anais, e outras informações das tabelas ESPÉCIES e STOCKS. Resulta
depois num relatório da informação diponível para a espécie juntamente com
todas as referências utilizadas (veja Relatórios, neste volume).
Fontes A tabela MORFOLOGIA, reúne dados de todos os catálogos
da FAO publicados até à data; de outras revisões taxonómicas; de livros de
faunas e de outras publicações periódicas, ex. Burges (1978), Trewavas (1983),
Allen (1985), Cohen et al. (1990), Lévêque (1990), Randall et al. (1990), Allen
(1991), Myers (1991), e Heemstra and Randall (1993).
Como Lá Chegar Chega-se à tabela MORFOLOGIA clicando no botão Biologia na janela ESPÉCIES, no botão Morfologia e Fisiologia na janela
BIOLOGIA e no botão Morfologia na
janela seguinte. Obtém-se o procedimento de identificação rápida clicando no
botão Espécies do Menu principal e
no botão Identificação rápida na
janela PROCURAR POR (...).
Referências Allen, G.R. 1985. FAO species catalogue. Vol. 6.
Snappers of the world. An annotated and illustrated catalogue of lutjanid
species known to date. FAO Fish. Synop. 6(125):208 p.
Allen, G.R. 1991. Damselfishes of the world. Mergus
Publishers, Melle, Germany. 271 p.
Burgess, W.E. 1978. Butterflyfishes of the world. A monograph
of the Family Chaetodonthidae. T.F.H. Publications, Neptune City. 832 p.
Cohen, D.M., T. Inada, T. Iwamoto and N.
Scialabba. 1990. FAO species catalogue. Gadiform fishes of the world
(Order Gadiformes). An annotated and illustrated catalogue of cods, hakes,
grenadiers and other gadiform fishes known to date. FAO Fish. Synop.
10(125):442 p.
Froese, R., and C. Papasisi. 1990. The use of modern relational databases for identification of fish larvae. J. Appl. Ichthyol. 6: 37-45.
Heemstra, P.C. and J.E. Randall. 1993. FAO species catalogue. Vol. 16. Groupers of the world.
(Family Serranidae, Subfamily Epinephelinae). An annotated and illustrated
catalogue of the grouper, rockcod, hind, coral grouper and lyretail species
known to date. FAO Fisheries Synop. 16(125): 382 p.
Lagler, K.F., J.E. Bardach, R.R. Miller, and D.R. May-Passino. 1977. Ichthyology. Second edition. John Wiley and
Sons, New York. 506 p.
Lévêque, C. 1990. Cyprinidae, p. 269-361. In C. Lévêque, D. Paugy and G.G. Teugels
(eds.) Faune des poissons d'eaux douces et saumâtres d'Afrique de l'Ouest. Tome
I. Coll. Faune Tropicale n° XXVIII. Musée Royal de l'Afrique Centrale,
Tervuren, and O.R.S.T.O.M., Paris. 384 p.
Myers, R.F. 1991. Micronesian reef fishes. Second edition.
Coral Graphics, Barrigada, Guam. 298 p.
Randall, J.E., G.R. Allen and R.C. Steene. 1990. Fishes of the Great Barrier Reef and Coral
Sea. University of Hawaii Press, Honolulu, Hawaii. 506 p.
Skelton, P.H. 1993. A
complete guide to the freshwater fishes of Southern Africa. Southern Book
Publishers, South Africa. 388 p.
Smith, M.M. and P.C. Heemstra, Editors. 1986. Smith’s sea fishes. Springer Verlag,
Berlin. 1047 p.
Trewavas, E. 1983. Tilapiine fishes of the Genera Sarotherodon, Oreochromis and Danakilia. British Museum of Natural
History, London, UK. 583 p.
Rainer Froese e Rodolfo B. Reyes, Jr.
Este
espaço dedicado à visão dos peixes, concentra-se no pigmento da retina do olho
do peixe e é baseado nos trabalhos de Denton e Warren (1956), Munz (1964), Munz
e McFarland (1973), Ali e Wagner (1975), e Hobson et al. (1981), dos quais,
foram extraídos até agora todos os registos (409), de 371 espécies.
A sensibilidade dos olhos dos peixes é máxima a um
certo comprimento de onda
Campos Os autores acima mencionados, mostraram que a sensibilidade do olho do peixe é máxima
a um certo comprimento de onda (em nm),
e o seu valor, é dado geralmente sob a forma de lmax (com
um intervalo de confiança de 95%), sendo este a chave de entrada desta tabela.
Um campo
sim/não permite registar a presença de outros pigmentos (como na Tabela 3 de
Hobson et al. 1981).
Um campo
de texto para notas completa esta pequena tabela.
Os
utilizadores desta tabela devem ler os artigos acima mencionados para detalhes
sobre os métodos utilizados para estimar lmax .
Estado Actualizar a tabela visão envolve:
·
incluir todas as
espécies descritas nos artigos acima citados (identificando os nomes válidos
das várias espécies, para as quais os autores utilizaram nomes não
actualizados);
·
adicionar novos
registos de artigos mais recentes, para serem identificados utilizando o Índice
de Citações científicas (através
da citação de alguma das três referências acima mencionadas); e
·
adicionar informação do
tamanho relativo do olho do peixe, assim como a sua distribuição batimétrica,
ambas relacionadas com o lmax.
A
informação desta tabela pode ser utilizada para testar as hipóteses
relacionadas com a fisiologia e ecologia dos peixes, tal como iniciado nas
referências abaixo mencionadas.
Como Lá Chegar Chega-se à tabela VISÃO clicando no botão Biologia na janela ESPÉCIES, e no botão
Pigmento do olho na janela BIOLOGIA.
Referências Ali, M.A. and H.J. Wagner. 1975. Visual pigments:
phylogeny and ecology, p. 481-516. In
M.A. Ali (ed.) Vision in fishes. New approaches to research. Plenum Press, New
York & London.
Denton, E.J. and F.J.
Warren. 1956. Visual pigments of deep-sea fish. Nature
4541:1059.
Hobson, E.S., W.N. McFarland and J.R. Chess. 1981. Crepuscular and nocturnal activities of
Californian nearshore fishes, with consideration of their scotopic visual
pigments and the photic environment. Fish. Bull. 79:1-30.
Munz, F.W. 1964. The visual pigments of epipelagic and
rocky-shore fishes. Vision Res. 4:441-454.
Munz, F.W. and W.N. McFarland. 1973. The significance of spectral position in the
rhodopsins of tropical marine fishes. Vision Res. 13:1829-1874.
Daniel Pauly
Os
peixes têm cérebros pequenos, pelo menos quando comparados com os vertebrados
superiores. No entanto, utilizar este facto contra os peixes é tão disparatado
como julgar o valor das pessoas apenas pelo tamanho dos cérebros (Gould, 1981).
O que
devemos fazer é apercebermo-nos de que o peixe tem o tamanho do cérebro que
precisa, e deste modo, a utilização das diferenças de tamanho de cérebro entre
as espécies, serve para induzir as diferentes necessidades de cada uma, i.e.,
no seu nicho (ver Bauchot et al. 1989). A base de dados “tamanho do cérebro”
composta por Roland Bauchot e seus colaboradores e gentilmente disponibilizada
para incluir nesta tabela da FishBase, permite-nos pensar desta forma. A seguir
descrevemos, baseado em Bauchot e Bauchot (1986), como é que esta base de dados
foi criada.
Mais de
2800 cérebros de cerca de 900 espécies de teleósteos foram dissecados (ver
Figura. 35). Muitos destes peixes foram recolhidos em localidades tropicais e
sub-tropicais tais como as Ilhas Havaianas e as Ilhas Marshall; Nova Caledónia;
Queensland, Austrália; Filipinas; Sudoeste da Índia; Maurícias e Reunião; Golfo
de Oman, a Norte do Mar Vermelho; Senegal e Caraíbas, mas também em França e no
Atlântico Norte. Todos estes peixes foram pesados e medidos (comprimento
standard ou total) antes da extracção dos cérebros. Os cérebros foram cortados
ao nível dos primeiros nervos espinhais, as meninges removidas, o cérebro pesado e posteriormente preservado em
Bouin.
Caixa 23.
Tamanho do cérebro e consumo de oxigénio.
Com uma grande série de dados disponível sobre o tamanho relativo do
cérebro, não resistimos a testar algumas hipóteses. A figura 36 mostra uma
tentativa de relacionar os dados da tabela CÉREBRO com outros dados de
fisiologia, neste caso os da tabela OXIGÉNIO. Ambas as séries de dados contêm
medidas individuais de peixes, que se encontram nos dois casos fortemente
correlacionadas com o peso.
Deste modo, foi utilizado o declive da relação
log-log de consumo de oxigénio vs
peso do corpo e peso relativo do cérebro vs
peso do corpo, respectivamente (para todos os dados disponíveis) para corrigir
os valores individuais da influência do peso do corpo.
Para o gráfico do tamanho do cérebro vs consumo de O2, obtivemos
então a média dos valores disponíveis para espécies com pelo menos três regisitos
do tamanho do cérebro e consumo de oxigénio. Apesar de alguma variância, a
figura 36 mostra que a hipótese de que cérebros maiores requerem mais oxigénio
e que são por isso mais comuns em peixes activos e com elevadas taxas
metabólicas não pode ser refutada. Esperamos que a variância seja menor uma vez
que a tabela OXIGÉNIO (veja neste volume) seja completamente verificada, e que
as suas próprias fontes de variância sejam identificadas.
Rainer
Froese and Daniel
Pauly
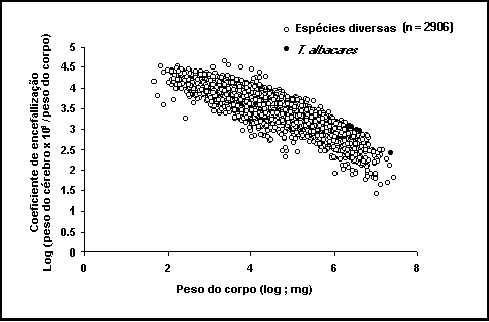
Fig. 35. Relação entre o peso relativo do cérebro e o
peso do corpo. Pontos claros: todos os registos da FishBase; pontos escuros:
dados para Thunnus albacares.
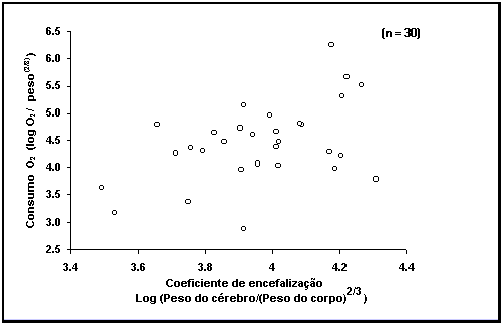
Fig. 36. Consumo de oxigénio vs peso relativo do cérebro em 30 espécies de peixes.
Os juvenis têm cérebros maiores
Como a proporção tamanho do
cérebro/peso do corpo é maior nos juvenis do que nos adultos (Bauchot et al.,
1979), utilizaram-se essencialmente adultos nas pesagens. No entanto, também
foram obtidas séries de dados, desde juvenis a grandes adultos, permitindo assim
um estudo ontogenético do tamanho do cérebro.
Os
registos obtidos para um peixe, estão aqui apresentados, com o nome corrente
das espécies, com os seguintes elementos:
·
peso do cérebro (em
mg);
·
peso do corpo (em g);
·
coeficiente de
encefalização (campo calculado= peso do corpo*1000/peso do corpo, cf Fig. 35); e
·
comprimento do corpo
(SL e/ou TL, em cm).
Os
registos são aqui apresentados, para cada espécie, sob a forma de tabela, com
pelo menos uma linha e até 73.
Futuramente,
esta tabela incorporará mais de 200 registos com os nomes das espécies, que até
agora foi impossível de relacionar com o nome válido da FishBase. Um dos nossos
elementos (James Albert) está a desenvolver esta tabela e já contribuiu com
registos de 77 espécies representando 18 novas famílias. Encontra-se em
preparação um artigo que analisa toda a série de dados (Albert e Froese, in
prep.)
Como chegar lá Clicando
no botão Biologia na janela ESPÉCIES, no botão Morfologia e Fisiologia na
janela BIOLOGIA e no botão Cérebro na janela seguinte.
Agradecimentos Queremos agradecer a R. Bauchot e aos seus
colaboradores terem confiado à FishBase a sua valiosa base de dados e a J. C.
Hureau por ter transformado esses dados num ficheiro legível pela equipa da
FishBase.
Referências
Bauchot,
R., M. Diagne and J.M. Ribet. 1979. Post-hatching growth and allometry of the
Teleost brain. J. Hirnforsch. 20:29-34.
Bauchot, M.L. and R. Bauchot. 1988. Encephalization in tropical teleost
fishes and its correlation with their locomotory habits, p. 678-690. In T. Uyeno, R. Arai, T. Taniuchi and K.
Matsuura (eds) Indo-Pacific Fish Biology: Proceedings of the Second
International Conference of Indo-Pacific Fishes. Ichthyological Society of
Japan, Tokyo.
Bauchot, R., J.M. Ridet and M-L. Bauchot. 1989. The brain organization
of butterflyfishes. Environ. Biol. Fishes 25 (1/3). 205-219.
Gould, S.J. 1981. The mismeasure of man. W.W. Norton, New York. 352 p.
Daniel Pauly
A informação sobre os requisitos de oxigénio é
importante na cultura de peixes
O metabolismo é um processo
fisiológico que reflecte o dispêndio de energia de um organismo vivo e por isso
os seus requisitos alimentares (nos heterotróficos). A taxa metabólica de um
peixe é geralmente medida pela sua taxa de respiração, i.e, a taxa de consumo
de oxigénio (Fig. 37). A informação sobre o consumo de oxigénio não só é útil em
fisiologia comparativa, como em cultura de peixes e gestão de pescas. Fornece
soluções de problemas associados com a rarefacção do peixe ou transporte de
peixe vivo, entre outros (Froese, 1988) (Caixa 22).
A tabela
OXIGÉNIO documenta o consumo de oxigénio de peixes, baseada em experiências
publicadas, juntamente com factores conhecidos ou sabidos que afectam a taxa
metabólica, tais como o sexo, peso do corpo, temperatura, salinidade,
concentração de oxigénio, nível de actividade e velocidade de natação e stress
aplicado. Detalhes adicionais de experiências, tais como número de peixes e
outras informações, encontram-se no campo comentários.
Os campos seguintes fornecem detalhes sobre os factores acima mencionados.
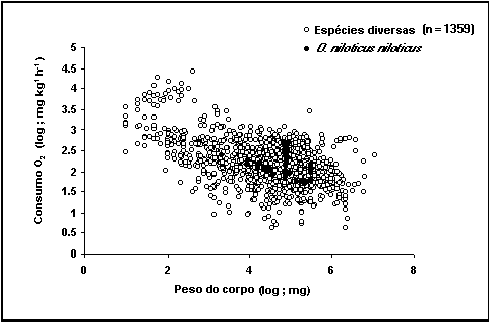
Fig. 37. Consumo relativo de oxigénio de Oreochromis niloticus niloticus
comparado com as restantes espécies. Repare no aumento causado pelo stress
aplicado para pesos compreendidos entre 100 e 300g.
Campos Consumo de
oxigénio: O consumo de oxigénio de
um peixe é dado em mg kg-1 h-1. Se o consumo foi apresentado noutra unidade, é
transformado em mg de oxigénio por kg de peixe por hora. Adicionalmente um
campo computorizado é incluído, no qual, o consumo de oxigénio a temperaturas
entre 5 e 30ºC é reexpresso no valor correspondente de consumo a 20ºC, o qual é
baseado na tabela 3.3 em Winberg (1971).
Sexo: É um
campo de escolha múltipla consistindo em: Fêmeas; machos; mistura(ambos os
sexos); sem sexo (para sexo desconhecido).
Peso:
Refere-se ao peso em gramas do organismo testado. Se a experiência foi
realizada com mais de um peixe, é utilizado o peso médio em gramas.
Número:
Refere-se ao número total de peixes utilizados na experiência.
Temperatura:
É a temperatura média da água, em ºC,
durante a experiência.
Salinidade: É
a salinidade média em permilagem durante a experiência. Se a salinidade não foi
indicada, assume-se que para espécies marinhas é de 35 por mil e 0 por mil para
espécies de água doce. Para peixes diádromos esta suposição é salientada no
campo Comentários.
Oxigénio (mmHg): Refere-se à pressão parcial do oxigénio na água da experiência em mmHg.
Como afirmado por Thurston e Gehrke (1993), este valor foi estimado fazendo
suposições baseadas na descrição dos métodos utilizados, quando este valor não
é dado na referência original. Estas suposições incluem correcções para a
temperatura e salinidade da experiência.
Oxigénio (mg/l): Este campo dá concentração de oxigénio da água da experiência em mg/l.
Se já existe uma entrada no campo de oxigénio (em mmHg) este valor não é retirado da literatura, mas calculado a
partir dos valores em mmHg, usando a
seguinte transformação:
![]()
Onde b é é o coeficiente de Bunsen para oxigénio a uma dada
temperatura e salinidade (Colt, 1984).
Saturação a 100% de oxigénio: Este campo existe, apenas para propósitos de
referência. Dá o oxigénio máximo calculado, em mg/l, da água a uma dada
temperatura e salinidade.
Variadas condições de stress foram aplicados aos
peixes
Percentagem de saturação: Este campo expressa, apenas como
referência, o conteúdo actual de oxigénio da água experimental, como percentagem
do máximo possível de oxigénio (ver stress aplicado).
Nível de actividade: É um campo de escolha que permite entrar em conta
com o efeito da actividade na taxa metabólica. As escolhas disponíveis para
este campo são: metabolismo padrão (peixe em descanso); metabolismo de rotina
(actividade espontânea do peixe); metabolismo activo (peixe em natação).
Velocidade de natação: Refere-se à velocidade de natação do peixe como
outro índice de actividade. A velocidade é representada como (ou convertida) em
comprimento do corpo por segundo (BL/s) com “B/L” correspondendo ao comprimento
total (TL) ou comprimento do corpo (FL).
Stress aplicado: É um campo de escolha que mostra o stress aplicado antes e durante a
experiência. As escolhas incluem: não especificado; temperatura (mudanças ou
valores extremos); fotoperíodo (duração invulgar ou tempo de exposição à luz);
alimentação (duração ou antes de experiência); jejum (supressão de comida por
mais de 24 horas); toxinas; hipoxia; hipercapnia (quantidade excessiva de dióxido
de carbono no sangue resultante de uma natação mais rápida forçada);
salinidade; valores elevados de pH; valores baixos de pH; sedativos;
transporte; outras formas de stress. Se a escolha for “outros”, o stress deve
estar especificado no campo comentários.
Utilizações A tabela OXIGÉNIO pode ser utilizada para: testar
hipóteses nas relações entre as diferentes actividades e stresses aos quais o
peixe é exposto; estimar consumo de energia (alimento) em modelos tróficos e
para relacionar crescimento, morfologia e taxas de metabolismo, entre outras
coisas.
A tabela OXIGÉNIO contém a maior colecção de dados
sobre o metabolismo dos peixes.
Fontes A tabela OXIGÉNIO contém provavelmente a maior
colecção de dados de consumo de oxigénio de peixes, com cerca de 7000 registos
de cerca de 300 espécies. A informação foi obtida de mais de 400 referências
tais como Neumann et al. (1981), Gorelova (1977), Marais (1978), Congleton
(1974), Winberg (1960), Subrahamanyam (1980). Destes dados, 6400 vêm da base de
dados “OXYREF” compilada por Thurston and Gehrke (1993), enquanto que o resto
foi adicionado pela equipa do FishBase.
Estado A verificação foi feita indo à literatura original e
verificando os valores e outra informação relevante aí registada. No entanto,
até à data, esta verificação foi apenas feita para algumas entradas. A equipa
da FishBase irá continuar a adicionar novos dados e a verificar a informação
entrada até agora.
Como Lá Chegar Chega-se à tabela OXIGÉNIO clicando no botão Biologia na janela ESPÉCIES, no botão Morfologia e Fisiologia na janela BIOLOGIA e no botão Metabolismo na janela seguinte.
Agradecimentos Estamos muito agradecidos a R. V. Thurston e P. C.
Gehrke pela oferta da base de dados OXYREF para distribuição através da
FishBase.
Referências Colt, J. 1984.
Computation of dissolved gas concentrations in water as functions of
temperature, salinity and pressure. Am. Fish. Soc. Spec. Publ. 14, 154 p.
Congleton, J.L. 1974. The respiratory response to asphyxia of Typhlogobius californiensis (Teleostei:
Gobiidae) and some related gobies. Biol. Bull. 146:186-205.
Froese, R. 1988. Relationship between body weight and loading
densities in fish transport using the plastic bag method. Aquacult. Fish.
Manage. 19:275-281.
Gorelova, T.A. 1977. Respiration rate of the fry of some
tropical fishes. Polskie Archiwum Hydrogiologii 24(Suppl.):447-453.
Marais, J.F.K. 1978. Routine oxygen consumption of Mugil
cephalus, Liza dumerii and L. richardsoni at different temperatures
and salinities. Mar. Biol. 50:9-16.
Neumann, D.A., J.M. O’Connor and J.A.
Sherk, Jr. 1981. Oxygen consumption of white perch (Morone americana), striped bass (M.
saxatilis) and spot (Leiostomus xanthurus).
Comp. Biochem. Physiol. 69A:467-478.
Subrahamanyam, C.B. 1980. Oxygen consumption of estuarine fish in
relation to external tension. Comp. Biochem. Physiol. 67A:129-133.
Thurston, R.V. and P.C. Gehrke. 1993. Respiratory oxygen requirements of fishes:
description of OXYREF, a data file based on test results reported in the
published literature, p. 95-108. In
R.C. Russo and R.V. Thurston (eds.) Fish physiology, toxicology, and water quality management. Proceedings of an International
Symposium, Sacramento, California, USA, 18-19 September 1990.
EPA/600/\R-93/157.
Winberg, G.G. 1960. Rate metabolism and food requirements of fishes. Translation Series No. 194.
Edited by F.E.J. Fry and W.E. Ricker. Fisheries Research Board of Canada, Biological
Station, Nanaimo, B.C.
Winberg, G.G. 1971. Methods for the estimation of production
of aquatic animals. Academic Press, London. 175 p.
Armi Torres e Rainer Froese
As tabelas
MODO DE NATAÇÃO e VELOCIDADE
A
imersão na água é a forma mais aproximada da não existência de peso. Para mais,
os teleósteos, através bexiga gasosa escapam ao efeito da força da gravidade
com o menor dispêndio de energia. O lado negativo de estar debaixo de água é o
facto de para cada movimento ter de deslocar água, um meio particularmente
denso e pesado. A física e fisiologia da natação dos peixes, está bem
sumarizada em Blake (1983) e Webb (1984), e a mais recente revisão neste campo
é dada por Videler (1993).
Esta
tabela descreve a forma na qual o peixe nada, o que é algo que faz num número
surpreendente de formas, e ainda os registos disponíveis de velocidade.
Os peixes nadam num número surpreendente de maneiras
A classificação aqui adoptada
provém de Lindsay (1978), que fez a revisão de muita da literatura antiga deste
tópico. Esta classificação consiste em dois níveis, o primeiro (números
romanos) descreve o que pode ser chamado o “tipo” de natação. O segundo (pontos
escuros) descreve o “modo” próprio de nadar.
I.
Movimentos do corpo e/ou barbatana caudal:
·
Anguiliforme
·
Subcarangiforme
·
Carangiforme
·
Tuniforme
·
Ostraciforme
II.
Ondulação das barbatanas médias ou peitorais:
·
Amiforme
·
Gimnotiforme
·
Balistiforme
·
Rajiforme
·
Diodontiforme
III.
Oscilações das barbatanas médias ou peitorais:
·
Tetraodontiforme
·
Labriforme
O modo
(I) implica uma transição gradual desde a ondulação do corpo inteiro (incluindo
o tronco) usado para propulsão (anguiliformes) até forças propulsivas
originadas apenas pela oscilação da barbatana caudal (modo tuniforme e
ostraciforme).
O leque
de formas em I, II, III, implica uma transição gradual de ondulações para
propulsões conforme os movimentos geram uma força propulsiva maior e por isso o
esquema acima indicado pode também ser representado em forma de gráfico, com um
eixo de ordenadas tronco-barbatana caudal e como abcissas ondulação-oscilação
(ver Figura 1 em Lindsay 1978).
Isto
implica que o modo de nadar de uma determinada espécie de peixe contém sempre
um elemento subjectivo mesmo ignorando o facto que o peixe pode ter dois modos
de nadar (considera-se aqui apenas o modo dominante, ex. labriforme no
papagaio-velho, fam. Scaridae, mesmo se ele reverter para o modo
subcarangiforme quando escapa de um presumível perigo (observ. pess. do autor).
Assim,
as entradas desta tabela (i.e., a escolha dos modos e tipos acima descritos),
pode ser revista de tempos a tempos, como resultado de uma familiarização com este
tópico e a sua literatura. No entanto, isto não afectará a primeira série de
entradas, baseadas nos exemplos de Lindsay e as suas extensões óbvias (ex. da Anguilla anguilla a todos os Anguilidae,
e destes a todos os anguiliformes). ( Espécies explicitamente dotadas de um
modo de nadar por Lindsay (1978) são diferenciadas de outras entradas na forma
Ref. 9574 (i.e. Lindsay 1978) nos campos Mainref
(referências principais) e Ref
(referências) ).
É de
notar também, que esta tabela apenas faz referência a peixes juvenis e adultos.
As larvas de peixe - por razões óbvias - têm um reportório limitado de tipos e
modos de natação.
O campo razão aspecto é definido como a razão
aspecto (A) da barbatana caudal,
calculado a partir de:
A= h2/ s
onde h é a altura da barbatana caudal e s é a área da sua superfície. O bordo
proximal da superfície da barbatana caudal é, por razões de melhor
reprodutibilidade, definida como uma linha direita vertical através da porção
mais próxima do pedúnculo caudal, i.e., considera-se a porção do pedúnculo
caudal rodeada pela barbatana como fazendo parte da área da barbatana caudal
(Fig. 38).
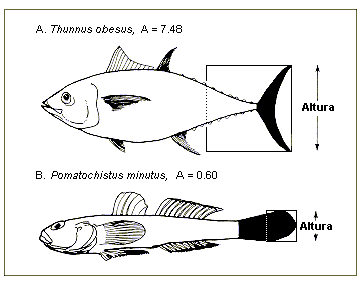
Fig.
38. Razão aspecto (A = h2/s, h = altura da barbatana caudal; s =
área de superfície da barbatana) de um peixe pelágico (A = 7.5) e de um peixe
bentónico (B = 0.6). Repare na correspondencia entre razões de aspecto e modos
de vida.
Repare
que esta definição de A difere
ligeiramente da apresentada por Pauly (1989), onde apenas a barbatana caudal é
utilizada para calcular s.
Pode ser dada uma referência para documentar uma razão aspecto. Quando
nenhuma referência é fornecida, a razão aspecto foi calculada por planimetria,
a partir de uma fotografia de um peixa na FishBase ou de outra fonte acessível.
Status
A tabela
MODO de NATAÇÃO contem o modo e tipo natatório para mais de 2600 espécies. R.W.
Blake, da Universidade de Colômbia britânica, ajudou-nos na verificação e
entrada de novos registos. A razão aspecto está diponível para mais de 115 espécies.
A tabela
VELOCIDADE, até à data contém mais de 240 registos com as velocidade máximas de
natação de 80 espécies. A informação foi extraída de mais de 50 referências
tais como Bainebridge (1958, 1960) e Webb (1971) e de compilações tais como
Sambilay (1990). Foi feito um esforço para distinguir natação sustentada (i.e.
mantida por mais de 3 minutos), e espontânea (mantida por apenas alguns
segundos) (Fig. 39), assim como outros modos de natação (Hammer, 1995).
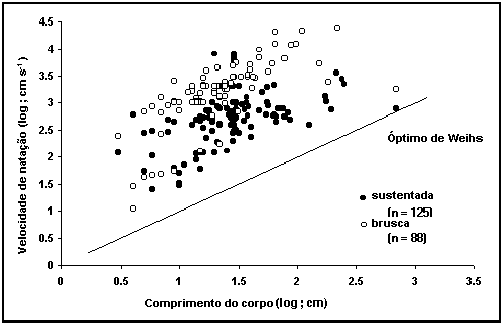
Fig.
39. Relação entre a velocidade e o comprimento do corpo. Repare que a
velocidade instantânea é de cerca de 10 vezes superior que a sustentada. Repare
ainda que nenhum peixe parece ter velocidades inferiores à linha de Weihs (1973) de energia mínima dispendida
por distância percorrida.
Campos A tabela VELOCIDADE consiste nos seguintes ramos:
Comprimento:
Este campo dá o comprimento do peixe como encontrado na publicação. O tipo de
medida do comprimento utilizado, consiste nas seguintes escolhas: SL
(comprimento padrão); FL (comprimento do corpo); TL (comprimento total); BL
(para o termo “comprimento do corpo” quando existe um comprimento mas o tipo de
medição não está indicado).
Velocidade de natação: Dá a velocidade do peixe por segundo, tal como
indicado na referência. Um campo de escolha é fornecido para o tipo de medida
de comprimento utilizada. As escolhas são as mesmas que no campo comprimento
(SL, FL, TL, BL) (metro por segundo). Um campo velocidade calculado em metro por segundo é também fornecido (ver valores derivados).
Modo: Este
campo descreve o modo de natação como: sustentado (nadando a uma velocidade que
dura um tempo prolongado); espontânea (velocidade máxima de natação que apenas
pode ser mantida menos de um minuto); outra.
Comentários:
Relata a medida de comprimento ou peso, ou modo de natação se estes não
estiverem na lista de escolha. Alguma transformação no tipo de comprimento
também deve ser aqui relatada.
Valores derivados: Dá os valores de transformação de diferentes unidades de velocidade
para comprimento padrão por segundo (SL/s) e para metro por segundo (m/s).
Torna assim possível comparar os desempenhos da natação de peixes com
barbatanas caudais em furcas, redondas, ou com outras formas.
Quando
disponível, os valores a e b usados para transformar comprimento
do corpo ou comprimento total em comprimento padrão são dados:
SL = a+b*L
onde L é o comprimento do corpo ou total
relatado. Se a transformação é apenas baseada numa medida de apenas um adulto
típico, a é zero.
Está
planeado rever e expandir os dados recolhidos até à data e John J. Videler
ofereceu-se gentilmente para ajudar neste assunto.
Como Lá Chegar Chega-se à tabela MODO DE NATAÇÃO E VELOCIDADE
clicando no botão Biologia na janela
ESPÉCIES, o botão Morfologia e
Fisiologia da janela BIOLOGIA, e o botão Velocidade De Natação ou Modo
de natação na janela seguinte.
Referências
Bainbridge, R. 1958. The
speed of swimming of fish as related to the size and to the frequency and
amplitude of the tail beat. J. Exp. Biol. 35(1):109-133.
Bainbridge, R. 1960. Speed
and stamina in three fish. J. Exp. Biol. 37(1):129-153.
Blake, R.W. 1983.
Functional design and burst-and-coast swimming in fishes. Can. J. Zool.
61:2491-2494.
Hammer, C. 1995. Fatigue
and exercise tests with fish. Comp. Biochem. Phsyiol. 112A(1):1-20.
Lindsey, C.C. 1978. Form,
function, and locomotory habits in fish, p. 1-100. In W.S. Hoar and D.J. Randall (eds.) Fish physiology. Vol. II.
Academic Press, New York.
Pauly, D. 1989. A
simple index of metabolic level in fishes. Fishbyte 7(1):22.
Sambilay, V.C., Jr. 1990.
Interrelationships between swimming speed, caudal fin aspect ratio and body
length of fishes. Fishbyte 8(3):16-20.
Videler, J.J. 1993. Fish
swimming. Chapman and Hall, London. 260 p.
Webb, P.W. 1971. The
swimming energetics of trout. Thrust and power output at cruising speed. J.
Exp. Biol. 55:489-520.
Webb, P.W. 1984. Form
and function in fish swimming. Sci. Am. 251:58-68.
Weihs, D. 1973. Optimal
fish cruising speed. Nature 245 (Sept. 7):48-50.
Rainer
Froese, Armi Torres, Cripina Binohlan e Daniel Pauly
Como
todos os animais heterotróficos, os peixes necessitam de alimento e oxigénio
para a sua sobrevivência, crecimento e reprodução. Contudo, apesar de existir
uma vasta literatura sobre alimento e hábitos alimentares dos peixes (incluida
nas tabelas e gráficos da FishBase),
existe muito menos informação sobre os orgãos e processos que permitem a
extração de energia destes alimentos.
O
processo-chave é a respiração, e está referido em parte na tabela OXIGÉNIO. Os
orgãos principais - as brânquias- são o tema da presente tabela.
Esta
tabela apresenta a maior parte das medições de área branquial de peixes,
publicadas até à data, i.e., medições da superfície que limita a entrada de
oxigénio, e por isso limita também a taxa de metabolismo e de crescimento
(Pauly 1979, 1981, 1994). A maior parte das informações provêm das compilações
de Hughes e Morgan (1973), Jagger e Dekkens (1985), e Palzenberger e Pohla
(1992).
Hughes
(1984) discute alguns dos problemas relacionados com a medição da superfície
branquial e com a sua interpretação. Esse trabalho deve ser consultado antes de
analisar a informação desta tabela. Pauly (1979, 1981, 1994) e Longhurst e
Pauly (1987) apresentam elementos sobre uma teoria de crescimento de peixes, na
qual a hipótese pode ser testada utilizando medições da superfície branquial.
Algumas utilizações práticas desta tabela incluem estudos de poluição e
ecotoxicologia.
A figura
40 mostra os registos de superfície branquial vs peso do corpo. Tal como esperado, esta relação bi-logarítmica
mostra uma diminuição da área branquial relativa com o peso do corpo, com um
declive de cerca da –0.2. Contudo, este gráfico esconde diferenças particulares
de cada espécie, que são importantes no estudo da relação entre a área
branquial e o crescimento (Pauly 1981).
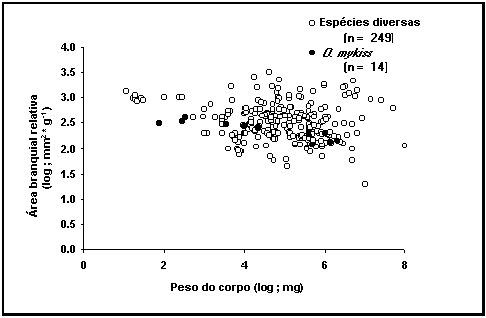
Fig. 40. Relação entre a superfície branquial de Oncorhynchus mykiss e o peso do corpo,
comparada com as restantes espécies. Pontos claros: todos os dados; Pontos
escuros: registos para a truta arco-íris.
Dadas
estas diferenças será provavelmente necessário considerar os modos natatórios e
e/ou a razão do aspecto da barbatana caudal.
Esperamos
ter na FishBase um gráfico que relacione directamente o crescimento e a
superfície branquial, tendo em conta estes factores.
O
conteúdo desta tabela irá sendo actualizado, utilizando referências apropriadas.
Qualquer sugestão da parte dos utilizadores sobre este assunto será bem-vinda.
Campos O campo chave desta tabela é superfície branquial (em cm2), que está sempre relacionada com o peso do corpo (em g).
Um campo
para a variável daí derivada, superfície
branquial/peso do corpo (em cm2/g), está também disponível, assim como um
campo para a distância sangue/água, i.e., espessura do epitélio branquial (em nm).
O campo Notas, permite comentários sobre a
metodologia, assim como outros comentários.
Como Lá Chegar Chega-se à tabela SUPERFÍCIE BRANQUIAL clicando no
botão Biologia na janela ESPÉCIES,
no botão Morfologia e Fisiologia na
janela BIOLOGIA, e no botão Superfície
Branquial na janela seguinte.
Agradecimentos Quero agradecer ao Professor G. M. Hughes a sua prontidão
de resposta, ao longo de todos estes anos, às minhas inúmeras questões sobre
brânquias de peixes.
Referências De Jager, S. and W. J.
Dekkers. 1975. Relation between gill structure and
activity
in fish. Neth. J. Zool. 25:276-308.
Hughes, G.M. 1984. Measurement of gill area in fishes: practices and
proble. J. Mar. Biol. Ass. U.K. 64:637-655.
Hughes, G.M. and M. Morgan. 1973. The structure of fish gills in
relation to their respiratory function. Biol. Rev. 48:419-475.
Longhurst, A. and D. Pauly. 1987. Ecology of tropical oceans. Academic
Press, San Diego, 407 p.
Palzenberger, M. and H. Pohla. 1992. Gill surface area of water
breathing freshwater fish. Rev. Fish. Biol. Fish. 2:187-216.
Pauly, D. 1979. Gill size and temperature as governing factors in fish
growth: a generalization of von Bertalanffy’s growth formula. Berichte des
Instituts für Meereskunde an der Universität, Kiel. No. 63, xv+ 156 p.
Pauly, D. 1981. The relationship between gill surface area and growth
performance in fish: a generalization of von Bertalanffy’s theory of growth.
Meeresforschung/Reports on Marine Research 28(4): 251-282.
Pauly, D. 1994. On the sex of fish and the gender of scientists. Chapman
and Hall, London, 250 p.
Daniel Pauly
Esta
tabela tenta incorporar na FishBase peixes como produto de consumo.
Campos Presentemente esta tabela consiste em 4 elementos:
1.
Uma série de campos que
as espécies e Stock, e a Localidade de onde as amostras de peixe
foram obtidas (pois existem diferenças regionais de sabor, textura, etc.)
2.
A FishBase como produto consumível
Campos que apresentam a
contribuição das diferentes partes do corpo (cabeça, tronco, pele, barbatanas, ossos, carne, filetes, vísceras, ovas, testículos, fígado) para
o peso total do peixe.
3.
Campos que apresentam a
composição química (percentagem de humidade,
proteína, gordura e cinzas) das
diferentes partes do corpo (carne/filetes; fígado; ovas; vísceras; cabeça/
ossos/ barbatanas; desperdício.); e
4.
O campo Notas que apresenta as propriedades
organolépticas dos diferentes modos de preparação (frito, fumado, enlatado,
etc.). Campos com Comentários são
também fornecidospara outras notas relacionadas com a proporção de peso e
composição química.
Fontes Quase todas as entradas até à data (680 registos de
500 espécies) provêm de Bykov (1983); de facto, a tabela PROCESSAMENTO foi
largamente desenvolvida para facilitar as entradas deste livro.
No
entanto, sentimos que esta tabela precisa de ser revista, tal como:
·
substituir as entradas
muito vagas no campo Notas, por uma
série de campos de escolha múltipla, o que permitirá uma padronização das
propriedades organolépticas dos modos de processar e da descrição física.
·
integrar as análises
químicas extensas e detalhadas de Vinogradov (1953), dos produtos da OECD
(1978) e de outras compilações semelhantes, e possivelmente de livros de
cozinha e receitas de peixe de várias culturas; e
·
um elo de ligação com a
tabela NOMES COMUNS, que também inclui (resume) descrições de produtos (ver
“tabela NOMES COMUNS”, neste volume).
Apreciaríamos
uma resposta dos colegas interessados em colaborar connosco neste assunto - um
desenvolvimento que tornará a FishBase útil para um grande número de novos
utilizadores.
Como Lá Chegar Chega-se à tabela PROCESSAMENTO clicando no botão Biologia na janela ESPÉCIES, no botão Peixes como consumo na janela BIOLOGIA,
e no botão Processamento na janela
seguinte.
Referências Bykov, V.P. 1983. Marine
Fishes: chemical composition and processing
properties.
Amerind Publishing Co., New Delhi.
OECD. 1978. Multilingual dictionary of fish and fish products.
Organization for Economic Co-operation and Development, Paris. 430 p.
Vinogradov, A.P. 1953. The elementary chemical composition of marine
organisms. Memoirs, Sears Foundation for Marine Research II, New Haven. 647 p.
Daniel Pauly, Emily Capuli e Rainer Froese