Esta tabela contém informação sobre crescimento, mortalidade natural e o comprimento até à primeira maturação, que são dados importantes na produção de modelos para a avaliação de stocks. Estes dados podem ainda ser utilizados na produção de relações empíricas entre os parâmetros de crescimento ou no cálculo de mortalidade natural e dados correlacionados (forma do corpo, temperatura, etc), que configuram uma linha de investigação útil para a avaliação dos stocks e também para conhecer melhor a evolução das estratégias biológicas (Fig. 16).
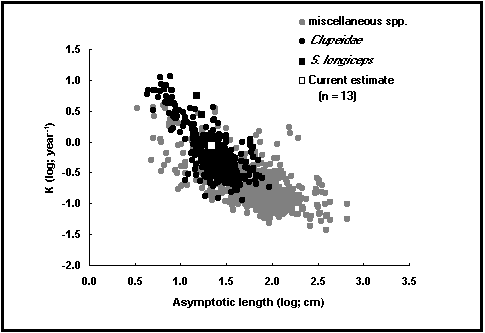
Fig. 16. Gráfico auxiométrico para Sardinella longiceps e para 20% dos dados de outras espécies.
Os parâmetros de crescimento incluídos nesta tabela são os da curva de von Bertalanffy (CVB; von Bertalanffy 1938), que têm a fórmula:
![]() ...1)
...1)
Onde Lt é o comprimento médio dum exemplar duma dada espécie com a idade t, L¥ é o seu comprimento médio assiptótico, isto é, o comprimento que atingiriam numa idade avançada, K é um factor de dimensão tempo-1, e to é a "idade teórica" (e geralmente negativa) que teria com o comprimento zero.
Em semelhança, o CVB para o crescimento em peso tem a fórmula:
Onde Wt e
W = a
A tabela POPCRESCIMENTO inclui registos para os quais, pelo menos, L¥ e K ou W¥ e K estão disponíveis, isto é, to pode não existir (este parâmetro não biológico não é utilizado na maior parte dos modelos de avaliação de stocks).
![]() ...2)
...2)
![]() ...3)
...3)
A tabela contém, presentemente, cerca de 4.700 conjuntos de parâmetros de crescimento calculados para mais de 1.150 espécies, extraídas de cerca de 2.000 fontes primárias e secundárias. As compilações de Pauly (1978, 1980) contribuíram com cerca de 1/3 das entradas.
Complementando a MainRef uma DataRef é dada para cada grupo de parâmetros de crescimento, porque frequentemente os parâmetros de crescimento são apresentados em artigos que não fornecem os dados de onde os resultados provêm. Os "dados da fonte", como indicado por um campo de escolha, consistem em: anéis dos otólitos, anéis das escamas, outros anéis anuais, anéis diários dos otólitos, marcação/recaptura, frequência de comprimento, observações directas, vários tipos de dados, outros.
Também o Método usado no cálculo é dado através da selecção num campo de escolhas com os seguintes itens: Ford-Walford; von Bartalanffy/Beverton; Gulland & Holt; regressão não linear; ELEFAN; outro(s) método(s).
A avaliação doutros métodos, as suas exigências, bias e outras necessidades dos dados podem ser consultados em Bugis (1976), Ricker (1980), Gulland (1983), Pauly (1984, 1997), Gayanilo & Pauly (1997) e noutros textos sobre haliêutica.
Para verificar o rigor dos parâmetros de crescimento incluímos:
um campo de cálculo com o índice de perfomance de crescimento Ø' =log10K +2log10L¥ ; Pauly 1979; Pauly & Munro, 1984 e ver também "Análises Auxiométricas", neste volume), que podem ser comparados com os valores de Ø'de outros stocks da mesma espécie ou de uma espécie estreitamente aparentada.
um campo de escolha múltipla descreve a forma como L¥ foi convertido em W¥, com as seguintes escolhas:
Tal como dado para crescimento em MainRef ou Ref.;
Calculado usando a relação L/W do mesmo stock;
Calculado usando a relação L/W de um stock diferente da mesma espécie;
Calculado utilizando a relação L/W de uma espécie similar;
Outros (ver Comentários)
um campo sim/não é usado para identificar os casos nos quais L¥ difere da Lmáx (na tabela ESPÉCIES) em mais de 30% de Lmáx;
quando estão disponíveis mais de 4 registos, um campo indica se o par de valores L¥- K se situa para lá da elipse auxiométrica (ver "Análises Auxiométricas", neste volume) definida pelos outros pares W¥-K para a espécie em questão.
um botão Curvas de crescimento gráfico na tabela resumo que, quando se clica, mostra um gráfico do tamanho do corpo vs idade relativa (Fig. 17) que pode ser utilizado para identificar curvas de crescimento que se desviem da tendência geral;
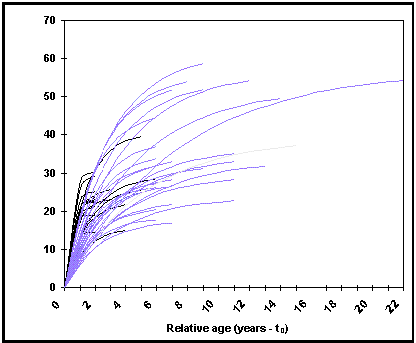
Fig. 17. Comprimento do corpo vs idade relativa (t-t0) em Oreochromis niloticus niloticus. As curvas foram calculadas aplicando a CVB (Equação 1) nos parâmentros L¥ e K da tabela POPCRESCIMENTO. As curvas de crescimento de pequena assímptota correspondem a crescimentos em cativeiro (ver Caixa 15 e Fig. 18).
botões de gráfico que mostram gráficos auxiométricos, por exemplo, plots de logK vs logL¥ (Fig. 16) e de W¥ (veja "Análises Auxiométricas", neste volume).
Um campo indica se o conjunto dos parâmetros de crescimento foram medidos em indivíduos selvagens ou cativos (ver Caixa15 e Figura 18).
As informações sobre tamanho na primeira maturação sexual (Lm), que aparece numa tabela separada (MATURAÇÃO), é usada em conjunção com L¥ para computar a "capacidade reprodutora" (Cushing, 1981) da população, ou seja, a razão Lm/L¥. A maior parte dos valores Lm referem-se ao comprimento médio ou ao comprimento a partir do qual 50% da população está sexualmente madura. Mas sempre que estes valores não são dados, ou não possam ser conseguidos a partir dos dados, Lm é o valor médio de dois valores limites.
Nas águas livres, as condições ambientais (por exemplo, a temperatura, mas também a presença de predadores), fazem comque os peixes cresçam lentamente até a um tamanho grande (K baixo, L¥ elevado), ou rapidamente até a um tamanho pequeno (K elevado, L¥ baixo). Por consequência, o índice de perfomance de crescimento (Ø'= log K + 2logL¥) fica quase constante entre as diferentes populações duma mesma espécie (Pauly 1994). As razões desta constância de Ø', que resulta em última análise do modo como os peixes distribuem a utilização do oxigénio difundido através da brânquias, são discutidas por Pauly (1981, 1994).
Para a maioria dos peixes que vivem em cativeiro, a ausência de predadores e de concorrentes sexuais permitem uma alocação do oxigénio mais importante para a alimentação e para o crescimento, visto que não o gastam em actividades de alto consumo de oxigénio, como a fuga aos predadores, ou no combate com os concorrentes sexuais.
Os índices Ø' dos indivíduos mantidos em cativeiro são por consequência superiores aos das populações selvagens. De facto, este efeito aumenta com a sofisticação do sistema de produção (Pauly et al. 1988). Evidentemente que este efeito será reforçado com o melhoramento genético do crescimento. Por exemplo, em Oreochromis niloticus (Tilápia-do-Nilo) (Pullin 1988) ou Salmo salar (Salmão-do-Atlântico) (Gjedrem 1985), a selecção dos indivíduos mais calmos, se bem que por vezes involuntária, favorece uma alocação óptima do oxigénio para o crescimento (Jones 1996; Bozynski 1988).
Destes efeitos combinados resultam valores de Ø' bem superiores para os indivíduos criados nos sistemas intensivos do que para os seus congéneres selvagens. Um gráfico foi desenvolvido para evidenciar estes efeitos, graças à adição dum campo na tabela POPCRESCIMENTO que distingue estes dois tipos de condiões de vida.
Como pode ser constatado na curva auxiométrica da figura 18, os pontos que se reportam aos peixes cativos desviam-se fortemente dos pontos referentes aos seus congéneres selvagens, sobretudo para os valores de L¥ compreendidos entre 10 e 30 cm que correspondem principalmente a Oreochromis niloticus criados em sistemas intensivos.
Referências
Bozynski, C. 1998. Growth, reproduction and behaviour of control and selected strains of Nile Tilapia (Oreochromis niloticus). Department of Zoology, University of British Columbia, M. Sc. Thesis.
Gjendrem, T. 1985. Improvement of productivity through breeding schemes. GeoJournal 10(3): 233-241.
Jones, R.E. 1996. Comparisom of some physical characteristics of salmonids under culture conditions using inderwater video imaging techniques. University of British Columbia, M. Sc. Thesis, 109 p.
Pauly, D. 1981. The relationships between gill surface area and growth perfomance in fish: a generalization of von Bertallanfy's theory of growth. Meersforschung 28(4): 251-282.
Pauly, D. 1994. On the sex of fish and the gender of scientists: essays in fisheries science. Chapman & Hall, London, 250 p.
Pauly, D., J. Moreau and M. Prein. 1988. Comparison of growth perfomance of tilapia in open water and aquaculture, p.469-479. In R.S.V. Pullin, T. Bhukaswan, K. Tonguthai and J.L. Mclean (ed.). Proceedings of the Second International Symposium on Tilapia in Aquaculture, 16-20 March 1987, Bangkok, Thailand. ICLARM Conf. Proc. 15.
Pullin, R.S.V., Editor. 1988. Tilapia genetics for aquaculture. ICLARM Conf. Proc.16, 108 p.
Daniel Pauly
Para alguns registos, as estimativas L ¥ têm ainda de ser confrontadas com os registos dos comprimentos máximos (Lmax) os quais, acreditamos, devem estar a priori razoavelmente próximos (ver acima).
Esperamos pelos comentários dos utilizadores acerca do conteúdo e/ou utilidade desta tabela.
Clique o botão Biologia na janela ESPÉCIES, o botão Dinâmica de populações na janela BIOLOGIA, e o botão Crescimento na janela DINÂMICA DE POPULAÇÕES. A figura 18 obtém-se também clicando o botão Relatórios na janela MENÚ PRINCIPAL seguido do botão Dinâmica das populações na janela MENÚ GRÁFICOS.
Bougis, P. Editor. 1976 Óceanographie biologique appliquée: l'exploitation de la vie marine. Masson, Paris, 320p.
Cushing, D.H. 1981. Fisheries biology: a study in population dynamics. 2nd ed. University of Wisconsin Press, Madison. 295 p.
Gayanilo, F.C. Jr., e D. Pauly. 1997. The FAO-ICLARM Stock Assessment Tool (FiSAT). Reference Manual. FAO Comp. Inf. Ser./Fish. 8, 262 p.
Gulland, J.A. 1983. Fish stock assessment: a manual of basic methods. FAO/Wiley, New York.
Pauly, D. 1978. A preliminary compilation of fish length growth parameters. Ber. Inst. Meereskd. an der Christian-Albrechts Univ. Kiel 55, 200 p.
Pauly, D. 1980. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J. Cons. CIEM 39(2):175-192.
Pauly, D. 1984. Fish population dynamics in tropical waters: a manual for use with programmable calculators. ICLARM Stud. Rev. 8, 325 p.
Pauly, D. 1997. Méthodes pour l'évaluation des ressources halieutiques. Collection Polytech. Cépaduès Editions, Toulouse, 288p.
Pauly, D. and J.L. Munro. 1984. Once more on growth comparison in fish and invertebrates. Fishbyte 2(1):21.
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 191. 382 p.
von Bertalanffy, L. 1938. A quantitative theory of organic laws II. Hum. Biol. 10:181-213.
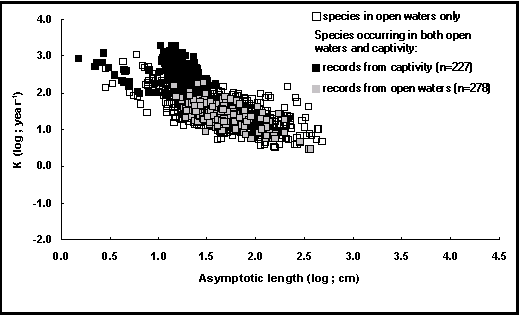
Fig.18. Nuvem auxiométrica visualizando o crescimento dos peixes em crescimento. Os quadrados negros em que o log(L¥) está compreendido entre 1,0 e 1,5 correspondem principalmente aos de Oreochromis niloticus criados em sistemas intensivos e semi-intensivos (ver Caixa 15).
Como chegar lá
Referências