A imersão na água é a forma mais aproximada da não existência de peso. Para mais, os teleósteos, através bexiga gasosa escapam ao efeito da força da gravidade com o menor dispêndio de energia. O lado negativo de estar debaixo de água é o facto de para cada movimento ter de deslocar água, um meio particularmente denso e pesado. A física e fisiologia da natação dos peixes, está bem sumarizada em Blake (1983), Webb (1984), e mais recentemente por Videler (1993).
Esta tabela descreve a forma na qual o peixe nada, o que é algo que faz num número surpreendente de formas, e ainda os registos disponíveis de velocidade.
A classificação aqui adoptada provém de Lindsay (1978), que fez a revisão de muita da literatura antiga deste tópico. Esta classificação consiste em dois níveis, o primeiro (números romanos) descreve o que pode ser chamado o "tipo" de natação. O segundo (pontos escuros) descreve o "modo" de nadar propriamente dito. viz.
Movimentos do corpo e/ou barbatana caudal:
Anguiliforme
Subcarangiforme
Carangiforme
Tuniforme
Ostraciforme
Ondulação das barbatanas médias ou peitorais:
Amiforme
Gimnotiforme
Balistiforme
Rajiforme
Diodontiforme
Oscilações das barbatanas médias ou peitorais:
Tetraodontiforme
Labriforme
O modo (I) implica uma transição gradual desde a ondulação do corpo inteiro (incluindo o tronco) usado para propulsão (anguiliformes) até forças propulsivas originadas apenas pela oscilação da barbatana caudal (modo tuniforme e ostraciforme).
O leque de formas em I, II, III, implica uma transição gradual de ondulações para propulsões conforme os movimentos geram uma força propulsiva maior e por isso o esquema acima indicado pode também ser representado em forma de gráfico, com um eixo de ordenadas tronco-barbatana caudal e como abcissas ondulação-oscilação (ver Figura 1 em Lindsay 1978).
A atribuição de um modo de natação a uma determinada espécie de peixe contém sempre um elemento subjectivo, mesmo ignorando o facto que o peixe pode ter dois modos de natação. Considera-se aqui apenas o modo dominante. Por exemplo os Scaridae passam de um modo labriforme em período normal para o modo subcarangiforme quando escapam de um presumível perigo (observ. pess. do autor).
Assim, as entradas desta tabela (i.e., a escolha dos modos e tipos acima descritos), pode ser revista de tempos a tempos, como resultado de uma familiarização com este tópico e a sua literatura. No entanto, os dados extraídos Lindsay (1978) (espécies que nos campos Mainref "referências principais" e Ref "referências" indiquem esta publicação) e as suas extensões óbvias (ex. da Anguilla anguilla a todos os Anguilidae, e destes a todos os anguiliformes) não serão afectadas por estas alterações.
É de notar também, que esta tabela apenas faz referência a peixes juvenis e adultos. As larvas de peixe - por razões óbvias - têm um repertório limitado de tipos e modos de natação.
O campo razão aspecto é definido como a razão aspecto (A) da barbatana caudal, calculado a partir de:
A = h2 / s
onde h é a altura da barbatana caudal e s é a área da sua superfície (Fig. 46). O bordo proximal da superfície da barbatana caudal é, por razões de melhor reprodutibilidade, definido como uma linha direita vertical através da porção mais próxima do pedúnculo caudal, i.e., considera-se a porção do pedúnculo caudal rodeada pela barbatana como fazendo parte da área da barbatana caudal.
Fig. 46. Razão aspecto (A = h2/s, h = altura da barbatana caudal; s = área de superfície da barbatana) de um peixe pelágico (A = 7.5) e de um peixe bentónico (B = 0.6). Repare na correspondência entre razões de aspecto e modos de vida. 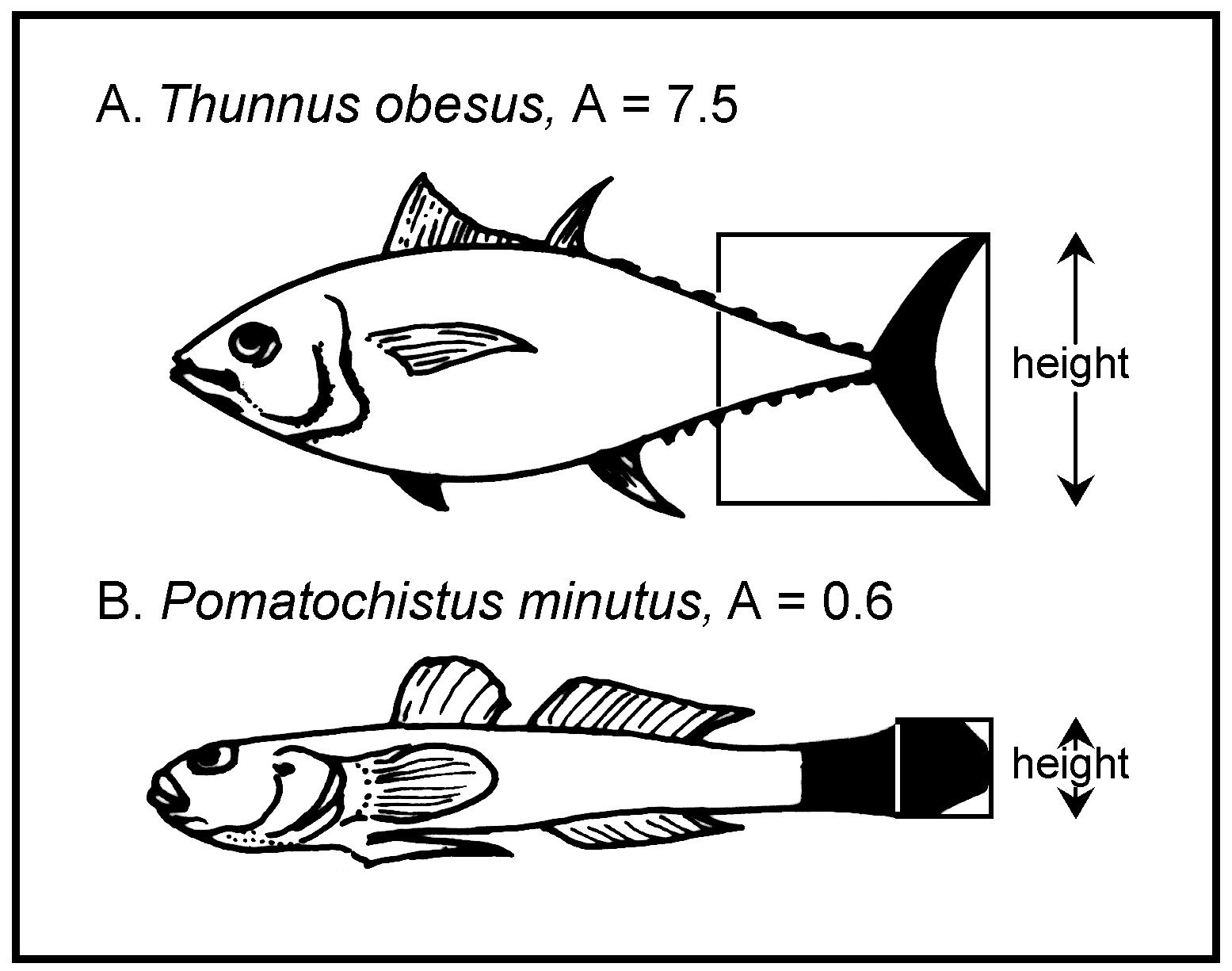
Repare que esta definição de A difere ligeiramente da apresentada por Pauly (1989), onde apenas a barbatana caudal é utilizada para calcular s.
Pode ser dada uma referência para documentar uma razão aspecto. Quando nenhuma referência é fornecida, a razão aspecto foi calculada por planimetria, a partir de uma fotografia de um peixe na FishBase ou de outra fonte acessível.
A tabela MODO de NATAÇÃO contem o modo e tipo natatório para mais de 2600 espécies.
A tabela VELOCIDADE, até à data contém mais de 247 registos com as velocidade máximas de natação de 79 espécies. A informação foi extraída de mais de 50 referências tais como Bainbridge (1958, 1960) e Webb (1971) e de compilações tais como Sambilay (1990). Foi feito um esforço para distinguir natação sustentada (i.e. mantida por mais de 3 minutos), e espontânea (mantida por apenas alguns segundos) (Fig. 47), assim como outros modos de natação (Hammer 1995).
A tabela VELOCIDADE consiste nos seguintes ramos:
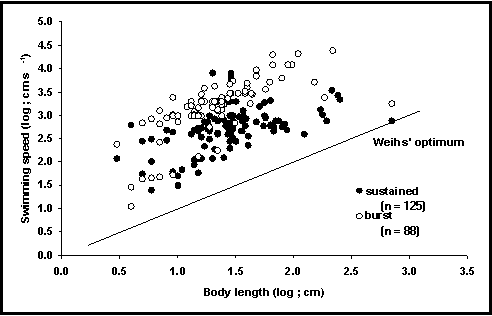
Fig. 47. Relação entre a velocidade e o comprimento do corpo. Repare que a velocidade instantânea é de cerca de 10 vezes superior que a sustentada. Repare ainda que nenhum peixe parece ter velocidades inferiores à linha de Weihs (1973) de energia mínima dispendida por distância percorrida.
Comprimento: Este campo dá o comprimento do peixe como encontrado na publicação. O tipo de medida do comprimento utilizado, consiste nas seguintes escolhas: SL (comprimento padrão); FL (comprimento do corpo); TL (comprimento total); BL (para o termo "comprimento do corpo" quando existe um comprimento mas o tipo de medição não está indicado).
Velocidade de natação: Dá a velocidade do peixe por segundo, tal como indicado na referência. Um campo de escolha é fornecido para o tipo de medida de comprimento utilizada. As escolhas são as mesmas que no campo comprimento (SL, FL, TL, BL) (metro por segundo).
Velocidade: Indica a velocidade convertida em metro por segundo (ver valores derivados).
Modo: Este campo descreve o modo de natação como: sustentado (nadando a uma velocidade que dura um tempo prolongado); espontânea (velocidade máxima de natação que apenas pode ser mantida menos de um minuto); outra.
Comentários: Relata a medida de comprimento ou peso, ou modo de natação se estes não estiverem na lista de escolha. Alguma transformação no tipo de comprimento também deve ser aqui relatada.
Valores derivados: Dá os valores de transformação de diferentes unidades de velocidade para comprimento padrão por segundo (SL/s) e para metro por segundo (m/s). Torna assim possível comparar os desempenhos da natação de peixes com barbatanas caudais em furcas, redondas, ou com outras formas. Quando disponível, os valores a e b usados para transformar comprimento do corpo ou comprimento total em comprimento padrão são dados:
LS = a + b × L
onde L é o comprimento do corpo ou total relatado. Se a transformação é apenas baseada numa medida de apenas um adulto típico, a é zero (ver a tabela COMPRIMENTO-COMPRIMENTO, neste volume).
Chega-se à tabela MODO DE NATAÇÃO E VELOCIDADE clicando no botão Biologia na tabela ESPÉCIES, o botão Morfologia e Fisiologia da tabela BIOLOGIA, e o botão Velocidade De Natação ou Modo de natação na janela MORFOLOGIA E FISIOLOGIA.
Bainbridge, R. 1958. The speed of swimming of fish as related to the size and to the frequency and amplitude of the tail beat. J. Exp. Biol. 35(1):109-133.
Bainbridge, R. 1960. Speed and stamina in three fish. J. Exp. Biol. 37(1):129-153.
Blake, R.W. 1983. Functional design and burst-and-coast swimming in fishes. Can. J. Zool. 61:2491-2494.
Hammer, C. 1995. Fatigue and exercise tests with fish. Comp. Biochem. Phsyiol. 112A(1):1-20.
Lindsey, C.C. 1978. Form, function, and locomotory habits in fish, p. 1-100. In W.S. Hoar and D.J. Randall (eds.) Fish physiology. Vol. II. Academic Press, New York.
Pauly, D. 1989. A simple index of metabolic level in fishes. Fishbyte 7(1):22.
Sambilay, V.C., Jr. 1990. Interrelationships between swimming speed, caudal fin aspect ratio and body length of fishes. Fishbyte 8(3):16-20.
Videler, J.J. 1993. Fish swimming. Chapman and Hall, London. 260 p.
Webb, P.W. 1971. The swimming energetics of trout. Thrust and power output at cruising speed. J. Exp. Biol. 55:489-520.
Webb, P.W. 1984. Form and function in fish swimming. Sci. Am. 251:58-68.
Weihs, D. 1973. Optimal fish cruising speed. Nature 245 (Sept. 7):48-50.
Rainer Froese, Armi Torres, Crispina Binohlan e Daniel Pauly